This Article
Creative Commons Attribution-NonCommercial 4.0 International License.
Serological Methods to Confirm Expression of Coat Protein Gene From an Iranian Isolate of Cucumber Mosaic Virus in Escherichia coli
Abstract
Background: Cucumber mosaic virus (CMV) has isometric particles with a diameter of about 28 – 29 nm. Detection and prevention are the critical steps in the control of plant viruses. Detection in a large number of samples is still done by serological methods due to their robustness and perhaps low cost.
Objectives: To this end, our aim was to express the CMV CP gene in E. coli to be used as the antigen for antibody production in the future.
Materials and Methods: Coat Protein (CP) gene cDNA from an isolate (B13) of Cucumber Mosaic Virus (CMV) was subcloned from pTZ57RCMVCP to pET21a expression vector and transformed to E. coli strain Rosetta. Expression of CMV CP was successful and confirmed by Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE), wherein a ~30- kDa protein band was revealed. Induction by Isopropyl-Thiogalactoside (IPTG) at final concentrations of 0.5 to 2 mM appeared to produce similar results as to the amount of the expressed protein, which was judged by intensity of the band on SDS-PAGE.
Results: The identity of the expressed protein was confirmed by immunoassays such as western blot, Dot-Immunobinding Assay (DIBA) and Enzyme-Linked Immunosorbent Assay (ELISA) by the use of anti-CMV antibody.
Conclusions: This is the first report of expression of CMV CP gene in Iran, which is important for the preparation of anti-CMV antibody and paving the way for the use of the virus coat protein as a nanomaterial.
Keywords: Methods; Cloning; Expression; Gene; Escherichia coli
1. Background
Cucumber Mosaic Virus (CMV) is a worldwide-spread plant virus with an extensive host range infecting about 1200 species in almost 85 families of plants including cereals, fruits, vegetables and ornamentals (1). It belongs to the genus cucumovirus in the family Bromoviridae. This virus causes symptoms such as yellow mosaic, leaf distortion and stunting on Nicotiana glutinosa. Cucumber Mosaic Virus particles are isometric with a diameter of about 28 – 29 nm (2, 3) and the virions consist of 180 identical copies of the virus-encoded coat protein (CP) (4). Cucumber Mosaic Virus is a multi- component virus consisting of three single-stranded genomic RNAs and two subgenomic RNAs known as sgRNA4 and sgRNA4A (2, 5, 6). Proteins encoded by RNAs 1 and 2 are needed for viral genome replication while RNA3 encodes the Movement Protein (MP) and Coat Protein (CP), both of which are involved in cell-to-cell and long-distance movement of the virus (2). The CMV CP is translated from subgenomic (sg) RNA 4 corresponding to the 3’ proximity of the viral RNA 3 (4). The sgRNA4A has been recognized as the viral repressor of host RNA silencing (VSR). The CMV is transmitted mechanically by sap and by aphids in a non-persistent, stylet-borne manner (7). Detection and prevention are the critical steps in the control of plant viruses. Detection in a large number of samples is still done by serological methods due to their robustness and perhaps low cost (8, 9). After the advent of recombinant DNA techniques, expression of viral genes in E. coli has been an important strategy for obtaining large-scale recombinant proteins that can be used for production of virus-specific antibodies (10–13).
2. Objectives
To this end, our aim was to express CMV CP gene in E. coli to be used as the antigen for antibody production in the future.
3. Materials and Methods
3.1. Cloning and Sequencing
The CMVCP gene was previously amplified from an Iranian CMV isolate (B13) by PCR, cloned in pTZ57R/T and sequenced with the following accession number AY871070 (13). Here, the CMV CP gene was subcloned from pTZ57CMVCP to pET21a (+) (Novagen, USA) by the use of BamHI and SacI sites (Figure 1). The recombinant plasmid pET21aCMVCP was used to transform the E. coli strain Rosetta by a heat shock procedure (14). Then, the plasmid was subjected to sequencing in order to ascertain the correct sequence before subjecting to expression (Macrogen, South Korea).
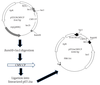 |
A Diagram of Subcloning of Cucumber Mosaic Virus Coat Protein Gene From pTZ57R/T Cloning Vector (left) into the pET21a Expression Vector (right)
|
3.2. Expression
A transformed colony of E. coli strain Rosetta carrying pET21aCMVCP was cultured overnight at 37°C in 5 mL of Luria Bertani (1% peptone, 0.5% yeast extract, 1% NaCl) containing 50 µg/mL ampicillin. Then, 1 mL of the overnight culture was added to 50 mL of fresh LB- ampicillin and growth was continued until the culture density reached an optical density of 0.6. Then, CP expression was induced by addition of Isopropyl-Thiogalactoside (IPTG) in the range of 0.5 to 2 mM (final concentration) and incubation at 37°C. Induction periods of 3 to 5 hours were applied. Then, the bacterial cells were harvested by centrifugation (5000 g at 4°C for 10 minutes) and stored at -80 °C for crude protein extraction.
3.3. Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis and Western Blotting
Proteins from the harvested cells were extracted with 160 μL of Laemmli buffer (125 mM Tris/HCl, pH 6.8, 2% sodium dodecyl sulfate, 9% glycerol, 0.7 M 2-mercaptoethanol, and 0.002% bromophenol blue) and boiled for five minutes. Proteins present in the supernatant and pellet fractions of the cell lysate were separated by 12% Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE) for four hours at 120 V (16) and visualized by 0.2% Coomassie Blue R staining solution. Then, the protein bands were electro blotted onto a HybondTM-N nitrocellulose membrane (Amersham, UK) using a wet transfer system (Akhtarian, Iran) at 100 mA during one hour for western blot analysis. Blocking was done by the use of 5% (w/v) Bovine serum albumin (BSA) powder in the buffer (8 mM Na2HPO4, 1.5 mM KH2PO4, 2.7mM KCl, 137 mM NaCl, pH 7.3) for one hour. After washing twice with Phosphate Buffered Saline-Tween 20 (PBST) membranes were incubated in 1:1000 dilution of polyclonal anti-CMV antibodies (Agdia, USA) for two hours at room temperature, followed by washing as above and incubating in alkaline phosphatase-conjugated anti-rabbit IgG for two hours at room temperature. After washing three times, the CMV CP specific band was visualized by incubating the membrane in the substrate Nitroblue Tetrazolium Chloride 5-Bromo-4-Chloro-3-Indolyl-Phosphate (NBT/BCIP), toluidine salt, Roche solution.
3.4. Dot Immunobinding Assay (DIBA)
The recombinant CMV CP extracted from E. coli was blotted onto a HybondTM-N nitrocellulose membrane. The membrane was dried at least for five minutes, immersed in PBST containing 1% BSA, and shaken and incubated for one hour at room temperature. Then, the membrane was washed three times with PBST (0.2 g KH2PO4, 2.9 g Na2HPO4.12H2O, 8g NaCl, 0.2 g KCl, 0.2 g NaN3, 1litre dH20 and 0.5 mL Tween 20) with agitation before incubation with the primary CMV-specific polyclonal antibody at a dilution of 1:1000 in PBST for two hours at 37°C. This was followed by washing of the membrane three times (as above), before incubating in conjugate anti-body (Alkaline Phosphatase-Labeled Goat Anti-Rabbit Globulin (APL -GAR)) diluted at 1:10000 in PBST for two hours at room temperature. Then, it was subjected to four washes as above. Finally, the membrane was incubated in NBT/BCIP substrate solution and then washed by distilled water to stop the reaction.
3.5. Enzyme-Linked Immunosorbent Assay
Enzyme-Linked Immunosorbent Assay (ELISA) was performed with the use of anti-CMV polyclonal antibody (Agdia, USA). Briefly, plates were coated with 100 μL (5 μg/mL) of antibody, incubated for three hours at 37°C, and washed three times with PBST. After blocking with PBST containing 1% BSA, 100 μL of the recombinant CMV CP extracted from 1.5 mL of bacterial culture was added, incubated for two hours at 37°C and washed by PBST (as above). Sap of CMV-infected cucumber leaves and bacterial cells carrying pET21a (no insert) were added as the positive and negative samples, respectively. Then, 1:2000 dilution of goat anti-rabbit IgG conjugate (Agdia, USA), was added and incubated for four hours at 37°C before washing five times with PBST. Finally, 150 μL of ρ-nitrophenyl phosphate solution (1 mg/mL) was added to each well and the yellow color development was monitored by the use of the Anthos 2020 (Austria) micro-titre ELISA reader by measuring absorbance values at 405 nm every 15 minutes for 90 minutes after the addition of the substrate.
4. Results
4.1. Preparation of Construct
Primers for the CMV CP cDNA amplification, which had been designed with restriction sites BamHI and SacI on the forward and reverse primer, respectively, facilitated in-frame cloning in the bacterial expression vector. After ligation of the CP cDNA into pET2la, subsequent sequencing confirmed correct insertion. As expected, additional nucleotides were inserted into the CP cDNA because of the restriction sites used for cloning; however, the insertion was in frame and no stop codon was created.
4.2. Expression of Coat Protein in Escherichia coli
Analysis by SDS-PAGE of total protein from bacterial cells that carried pET21aCMVCP and were induced by IPTG in the range of 0.5 to 2 mM (final concentration) for four hours showed a higher level of expression of the expected protein, approximately 25 kDa in size, corresponding to the CMV CP (17) (Figure 2A). Four independent transformants (clones) were examined for the expression, among which two clones with a desired level of the expression were selected for further analysis.
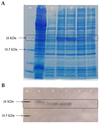 |
Analyses of Cucumber Mosaic Virus Coat Protein Expression by Examination of Protein from Escherichia coli With Different Methods
|
4.3. Detection by Western Blotting, Dot Immunobinding Assay and Enzyme-Linked Immunosorbent Assay
Western blotting and DIBA with a rabbit anti-CMV polyclonal antibody confirmed the identity of the recombinant protein as CMV CP. Accordingly, a protein band with 26 KDa molecular weight, which was the expected size (based on comparison with the protein size marker) was observed in the western blot (Figure 2B). In the DIBA, sap of CMV-infected cucumber leaf, which was used as a positive control, produced a signal after 15 to 30 minutesof incubation with the substrate (Figure 3A). While no reaction occurred with the extract from bacterial cells containing pET21a, only a weak reaction appeared with the non-induced bacteria (negative controls). Subjection to ELISA of sap from CMV-infected cucumber (positive control), and total protein from bacteria carrying the expression construct, pET21aCMVCP, resulted in production of yellow color 30 minutes after addition of the substrate whereas no signal was visually detected with the protein from bacterial cells containing pET21a(+) (no insert), which were used as the negative control (Figure 3A, 3B).
 |
Analysis of Recombinant Cucumber Mosaic Virus Coat Protein by the Use of Serological Methods
|
5. Discussion
All of the expression systems have advantages and limitations. E. coli as a prokaryotic system because of its high expression capacities of many heterologous proteins, low cost, efficient generation time and fast high-density cultivation is known as a successful host. Accordingly, expression of CP gene from numerous plant viruses has beensuccessfully done in E. coli (18–21). Apart from the pET vector system, a number of other vector systems such as pGEX (22, 23), pTrcHis (24) and pTBG (H) (25) have also been used for expression of foreign genes in E. coli. With all the vector systems available, E. coli strain Rosetta has been the cell line of choice for expression of genes of interest (26, 27). Strain Rosetta belongs to the so-called “B strains” that are deficient in lon protease and lack the ompT outer membrane protease. The strains having these proteases can degrade proteins during purification (28). Therefore, some target proteins may be more stable in these strains than in host strains containing these proteases. Also, Rosetta is an optimal strain when the target gene contains rare codons or when the expressed protein has disulfide bonds (29). The identity of the expressed protein was demonstrated to be CMV CP by western blotting, DIBA and ELISA. Although with the E. coli carrying pET21a (with the CP insert but not induced by IPTG) a weak reaction was obtained in western blotting and DIBA (Figure 2B, 3A), such background reactions are not unexpected considering the fact that a basal expression can occur in the absence of IPTG (29). Various IPTG concentrations have been used to induce expression of different genes of interest. These include IPTG at final concentration of 1 mM (25, 26, 30) and 0.4 mM (27) for expression of Cardamom mosaic virus CP and 0.1 mM (31) for expression of Prunus Necrotic Ring Spot Virus (PNRSV) CP in E. coli. In our study, we tested IPTG concentrations ranging from 0.5 to 2 mM and found that changes in this range do not affect expression level (data not shown). It has been reported that the duration for expression of gene of interest at a particular IPTG concentration may vary according to the gene of interest or other factors; for instance incubation at 37°C for three to four hours (26, 27, 32), and at 16°C overnight has been reported (30). In the present study, we used induction periods of three to five hours and found that an induction period of four hours gives the best result (data not shown). In our study, expression of the CMV CP gene under optimized conditions favored the expressed protein to be mostly present in the soluble fraction of the cell lysate. In most cases of heterologous expression of genes in E. coli, it has been reported that the expressed protein is present as an insoluble fraction at 37°C; however, when large-sized proteins are expressed in E. coli or when small-sized proteins are over-expressed, insoluble difficult-to-hydrolyze protein complexes are formed (33). This research showed that CMV CP open reading frame placed under the control of T7 promoter is expressed in E. coli. After induction of expression, the levels of CP were monitored over a four to five-hour period by SDS-PAGE and expression was confirmed by immunoassay tests including western blot, DIBA and ELISA.
Acknowledgments
We would like to acknowledge the Iranian National Science Foundation for providing funds to carry out this research.
Footnotes
References
- 1. Roossinck MJ. Cucumber mosaic virus, a model for RNA virus evolution. Mol Plant Pathol. 2001;2(2):59-63. [PubMed]
- 2. Palukaitis P, Roossinck MJ, Dietzgen RG, Francki RIB. Cucumber MOSAIC Virus. 1992;41:281-348.
- 3. Madhubala R, Bhadramurthy V, Bhat AI, Hareesh PS, Retheesh ST, Bhai RS. Occurrence of Cucumber mosaic virus on vanilla (Vanilla planifolia Andrews) in India. J Biosci. 2005;30(3):339-50. [PubMed]
- 4. Smith TJ, Chase E, Schmidt T, Perry KL. The structure of cucumber mosaic virus and comparison to cowpea chlorotic mottle virus. J Virol. 2000;74(16):7578-86. [PubMed]
- 5. Palukaitis P, García-Arenal F. Cucumoviruses. 2003;62:241-323.
- 6. Van Regenmortel MHV, Fauquet CM, Bishop DHL, Carstens EB, Estes MK, Lemon SM. Virus taxonomy: classification and nomenclature of viruses. Seventh report of the International Committee on Taxonomy of Viruses Seventh report of the International Committee on Taxonomy of Viruses. Academic Press; 2000.
- 7. Ng JC, Liu S, Perry KL. Cucumber mosaic virus mutants with altered physical properties and defective in aphid vector transmission. Virology. 2000;276(2):395-403. [PubMed]
- 8. Ling KS, Zhu HY, Jiang ZY, Gonsalves D. Effective application of DAS-ELISA for detection of grapevine leafroll associated closterovirus-3 using a polyclonal antiserum developed from recombinant coat protein. Eur J Plant Pathol. 2000;106(4):301-9.
- 9. Zimmermann D, Bass P, Legin R, Walter B. Characterization and Serological Detection of Four Closterovirus-like Particles Associated with Leafroll Disease on Grapevine. J Phytopathol. 1990;130(3):205-18.
- 10. Barbieri MR, Carvalho M, Zambolim EM, Zerbini FM. Expression in Escherichia coli of the capsid protein of Watermelon mosaic virus and production of specific antiserum. Phytopathol Braz . 2004;29(2):215-9.
- 11. Nickel O, Targon MLPN, Fajardo TVM, Machado MA, Trivilin AP. Polyclonal antibodies to the coat protein of Apple stem grooving virus expressed in Escherichia coli: production and use in immunodiagnosis. Phytopathol Braz . 2004;29(5):558-62.
- 12. Targon MLPN, Nikolaeva O, Manjunath KL, Lee RF, Mueller GW, Machado MA. Coat protein gene of a Brazilian isolate of the Citrus tristeza virus: cloning, expression in E. coli and production of polyclonal antiserum. Phytopathol Braz . 1997;22:99-102.
- 13. Bashir NS, Kalhor MR, Zarghani SN. Detection, differentiation and phylogenetic analysis of cucumber mosaic virus isolates from cucurbits in the northwest region of Iran. Virus Genes. 2006;32(3):277-88. [PubMed]
- 14. Chung CT, Niemela SL, Miller RH. One-step preparation of competent Escherichia coli: transformation and storage of bacterial cells in the same solution. Proceeding Natl Acad Sci. 1989;86(7):2172-5.
- 15. Kivirauma T, Oikari P, Saareta J. Plasmid Processor Version 1. Kuopio Finalnd: University of Kuopio; 1996.
- 16. Laemmli UK. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 1970;227(5259):680-5. [PubMed]
- 17. Natilla A, Hammond RW, Nemchinov LG. Epitope presentation system based on cucumber mosaic virus coat protein expressed from a potato virus X-based vector. Arch Virol. 2006;151(7):1373-86. [PubMed]
- 18. Abou-Jawdah Y, Sobh H, Cordahi N, Kawtharani H, Nemer G, Maxwell DP, et al. Immunodiagnosis of Prune dwarf virus using antiserum produced to its recombinant coat protein. J Virol Methods. 2004;121(1):31-8. [PubMed]
- 19. Chen CC, Hsiang T, Chiang FL, Chang CA. Molecular characterization of Tuberose mild mosaic virus and preparation of its antiserum to the coat protein expressed in bacteria. Bot Bull Academia Sin. 2002;43
- 20. Fajardo TVM, Barros DR, Nickel O, Kuhn GB, Zerbini FM. Expression of Grapevine leafroll-associated virus 3 coat protein gene in Escherichia coli and production of polyclonal antibodies. Phytopathol Braz . 2007;32(6):496-500.
- 21. Plchova H, Moravec T, Dedic P, Cerovska N. Expression of Recombinant Potato leafroll virus Structural and Non‐structural Proteins for Antibody Production. J Phytopathol. 2011;159(2):130-2.
- 22. Bicka L, Kuźmak J, Kozaczynska B, Plucienniczak A, Skorupska A. Expression of bovine leukemia virus protein p24 in Escherichia coli and its use in the immunoblotting assay. Acta Biochim Pol. 2000;48(1):227-32.
- 23. Hema M, Subba Reddy Ch V, Savithri HS, Sreenivasulu P. Assembly of recombinant coat protein of sugarcane streak mosaic virus into potyvirus-like particles. Indian J Exp Biol. 2008;46(11):793-6. [PubMed]
- 24. Hammond RW, Crosslin JM. The complete nucleotide sequence of RNA 3 of a peach isolate of Prunus necrotic ringspot virus. Virology. 1995;208(1):349-53. [PubMed]
- 25. Thomas JG, Baneyx F. Protein misfolding and inclusion body formation in recombinant Escherichia coli cells overexpressing Heat-shock proteins. J Biol Chem. 1996;271(19):11141-7. [PubMed]
- 26. Bragard C, Duncan GH, Wesley SV, Naidu RA, Mayo MA. Virus-like particles assemble in plants and bacteria expressing the coat protein gene of Indian peanut clump virus. J Gen Virol. 2000;81(Pt 1):267-72. [PubMed]
- 27. Jacob T, Usha R. Expression of Cardamom mosaic virus coat protein in Escherichia coli and its assembly into filamentous aggregates. Virus Res. 2002;86(1-2):133-41. [PubMed]
- 28. Grodberg J, Dunn JJ. ompT encodes the Escherichia coli outer membrane protease that cleaves T7 RNA polymerase during purification. J Bacteriol. 1988;170(3):1245-53. [PubMed]
- 29. Novagen EMD Biosciences. pET System Manual. 11th ed. Gibbstown; 2006.
- 30. Kadkhodayan S, Knapp MS, Schmidt JJ, Fabes SE, Rupp B, Balhorn R. Cloning, expression, and one-step purification of the minimal essential domain of the light chain of botulinum neurotoxin type A. Protein Expr Purif. 2000;19(1):125-30. [PubMed]
- 31. Petrzik K, Mraz I, Kubelkova D. Preparation of recombinant coat protein of Prunus necrotic ringspot virus. Acta Virol. 2001;45(1):61-3. [PubMed]
- 32. Liu W, He R, Xue Z. Expression of the n-terminal segment of qbrn-2 in E. coli and tips on preparation of a recombinant protein. Protein Pept Lett. 2001;8(1):27-32.
- 33. Koschorreck M, Fischer M, Barth S, Pleiss J. How to find soluble proteins: a comprehensive analysis of alpha/beta hydrolases for recombinant expression in E. coli. BMC Genomics. 2005;6:49. [PubMed]

